packz ha letto “Il gene egoista”
Posted: Febbraio 23rd, 2010 | Author: packz | Filed under: Letture | 2 Comments »
Ho finito di leggere "il gene egoista" di Dawkins, un saggio che tratta la visione "moderna" sulla teoria dell’evoluzione come si è andata affermando in questi ultimi anni, benché il volume sia uscito originariamente nel 1976; viene diviso in capitoli che idealmente seguono un percorso "evolutivo", dalla nascita della vita sino allo sviluppo di un nuovo brodo primordiale costituito dalla cultura umana ed i suoi nuovi replicatori; anche se mi sarebbe piaciuto suddividere in maniera simile la recensione sarebbe stata alla fine una scopiazzatura fin troppo palese del testo, invece riassumerò i punti (per me) cruciali usando anche parti di testo direttamente da Hofstader e da quel libro incredibile che è Metamagical Themas.
Iniziamo quindi (sperando di non dimostrare solo una inettitudine nello scrivere come temo)
Il gene egoista
Il libro inizia affermando che la risposta alla domanda "perché esiste la gente" non ha senso prima del 1859, anno in cui Darwin presenta la sua teoria dell’evoluzione: secondo questa teoria la diversità delle specie viventi è spiegabile attraverso un processo chiamato evoluzione, guidato in maniera attiva dalla cosiddetta selezione naturale che permette, in poche parole, solo al più adatto di sopravvivere. In questo libro si cerca di cambiare punto di vista e capire chi sia il vero attore del palcoscenico della vita biologica.
La storia inizia affrontando il problema della nascita della vita; per capire come può essere avvenuto questo evento, partiamo osservando che in natura esiste una legge più generale della legge di selezione naturale di Darwin, la legge della "stabilità": ogni aggregato che sia stabile per un certo periodo di tempo e abbastanza grosso, merita un nome a sè stante. Quando la terra si è formata, l’ambiente era molto diverso da come è attualmente, erano presenti specie chimiche non adatte alla vita a come la conosciamo oggi, è presumibile che siano state presenti quantità di ammoniaca, metano e idrogeno che andarono a formare il brodo primordiale; nell’esperimento di Miller-Urey è stato dimostrato che tali composti, se sottoposti a scariche elettriche o a radiazione (la fascia di Ozono è stata creata dalle specie viventi ;-)), sono capaci di formare delle sostanze chiamate aminoacidi, le quali sono alla base della vita.

Proprio alla luce di queste considerazioni si capisce il perché fa spesso notizia il trovare acqua (o tracce di essa) su un pianeta: è probabile che la vita come la conosciamo noi si sia sviluppata su di esso.
Ritornando agli eventi che noi interessano, è probabile che tra le migliaia di molecole che andarono a formare il contenuto del brodo se ne generò una particolare, capace di replicarsi, meritevole di un nome tutto suo: il replicatore; Dawkins ammette che questo processo non è probabile, ma neanche impossibile: una volta prodotta una singola unità capace di fare questo, essa si "riprodurrà" a spese del contenuto del brodo. Una molecola capace di riprodurre se stessa non è così difficile da immaginare, pensate ad una molecola capace di creare come uno stampo delle proprie copie o (come avviene nel DNA) dei negativi della copia. Una volta iniziato questo processo, si avrà una competizione fra le varie copie per lo sfruttamento delle molecole che dovevano utilizzare per replicarsi; ovviamente il processo di copiatura non sarà perfetto: a poco a poco una diffusione di replicatori aventi degli "errori" tali da renderli più stabili, più veloci (nell’eseguire una propria copia) causerebbe una loro tautologica selezione.
In effetti (questo è un mio pensiero) non è improbabile pensare che i ribosomi (gli organuli che si preoccupano di creare le proteine a partire dai codici di DNA) siano in realtà molecole createsi indipendentemente nel brodo primordiale (o in una fase successiva) e che avevano la proprietà di creare proteine a partire dai codici casuali nel brodo. Ovviamente è indubbio che pezzi di replicatori capaci di creare una membrana attorno ad essi avrebbero avuto un vantaggio rispetto a quelli non dotati di questo optional, senza contare il vantaggio di avere il ribosoma encodato dentro il replicatore; proprio a sostegno di questa ipotesi si ha la conoscenza che i mitocondri (altri organuli preposti alla gestione energetica delle cellule) utilizzano un proprio dialetto di DNA che si replica sono attraverso la generazione femminile (da qui alcune malattie solo femminili e la possibilità di studiare chi era Eva).È anche possibile che in realtà questi organuli non siano altro che ex parassiti delle cellule primordiali.
La competizione può aver portato alla crezione di una nuova dinastia di replicatori tali da scalzare il protagonista iniziale di questa storia, infatti il codice replicante attuale si trova encodato nel DNA ma non è detto che esso rappresenti il replicatore protagonista dell’inizio della storia, può essere che il nostro acido desossiribonucleico abbia scalzato il replicatore durante la storia per divenire il governante assoluto ai tempi nostri.
Tuttavia il protagonista dell’evoluzione non è il replicatore nella sua completezza, ma la sua unità più piccola: il gene. Per capire cosa si intenda per gene, Dawkins fa un piccolo escursus sulle caratteristiche del DNA che riassumo per quelli che non sono andati alle elementari: in pratica il DNA non è altro che una molecola complessa con una struttura ad elica, i cui componeti fondamentali sono i nucleotidi, cioè 4 molecole indicate con A, T, C, G le quali rappresentano l’alfabeto tramite cui "è scritta la vita". Queste sequenze di quattro lettere encodano, presi a gruppi di tre, degli aminoacidi specifici; queste sequenze di aminoacidi formano a loro volta delle proteine che vengono usate dalla cellula per i propri scopi.
A prescindere dal fenomeno di produzione di proteine, le lunghe eliche di DNA sono organizzate in cromosomi: se i geni rappresentano le pagine, dice Dawkins, allora i cromosomi rappresentano dei libri; la particolarità è che i cromosomi girano in coppie, ed il numero di coppie è variabile tra le specie (non è neanche detto che un numero maggiore di cromosomi indichi maggiore evoluzione, infatti una salamandra ha circa 20 volte i geni dell’essere umano). Sono in coppie perché un individuo eredita un elemento da ognuno dei genitori (ovviamente nelle specie sessuate); ognuno dei geni presenti nei cromosomi che occupa una specifica posizione, determina il cosidetto fenotipo (cioé il carattere esteriore "finale" prodotto da cotale gene) che tuttavia può differire tra due stessi geni occupanti la medesima posizione in una delle coppie dello stesso cromosoma (questi geni si chiamano alleli); come fa a decidere il corpo quale carattere presentare se il padre ha gli occhi verdi e la madre blu? se un allele domina sul rivale è detto dominante, altrimenti recessivo (ovviamente ci sono casi in cui entrambi concorrono nella formazione del fenotipo ma adesso non me ne vengono in mente). Le leggi che governano questo fenomeno furono scoperte da Mendel, un prete che fece i primi esperimenti di ereditarietà sulle piante di piselli. È interessante come la genetica nasca proprio dagli studi di un prete.
Per capire più o meno di cosa stiamo parlando ecco un bel disegnino di diffusioni dei geni in un sistema di riproduzione XY usato prevalentemente dai mammiferi (grazie alla mie possenti capacità di informarmi sono venuto a sapere che esistono molti modi diversi in natura oltre questo per determinare il sesso).
Le lettere sotto i simboli rappresentano le coppie di alleli presenti in ogni individuo
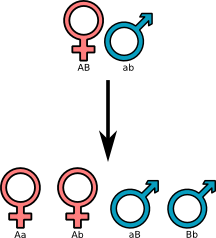
Come potete osservare ogni allele ha le stesse probabilità di presentarsi in un discendente. Dawkins indica con "lotteria meiotica" questa caratteristica e sottolinea che conviene a tutti i geni che sia così: se, facciamo un esempio, il cromosoma Y trovasse una maniera di riuscire a far produrre gameti contenenti solo esso al loro interno, avrebbe un vantaggio di diffusione rispetto agli altri ma porterebbe presto alla scomparsa della specie in quanto produrrebbe solo maschi; l’esempio tuttavia non è proprio campato in aria in quanto, relativamente al cromosoma X, esiste il fenomeno della lyonization che spiega anche il mantello del vostro gatto. In pratica è come se gli altri geni presenti nei cromosomi restanti si "mettessero d’accordo" per cancellare questo cromosoma.
Tuttavia anche se abbiamo parlato di geni fino ad adesso, la loro definizione è molto più sfumata; per averne una più precisa è necessario tirare di mezzo anche il crossing over: in pratica durante il processo della divisione cellulare delle cellule sessuali (per gli amici meiosi) le coppie di cromosomi si dividono e vanno ognuno in una cellula sessuale (per capirci lo spermatozoo e l’ovulo) formando così delle cellule aploidi; il fatto è che non si dividono e basta, ma eseguono una pratica chiamata appunto crossing over: si scambiano fra loro del materiale genetico, eseguendo un rimescolamento delle sequenze ereditarie. Capite che in questa maniera, in ogni nuova generazione, un cromosoma non è mai ereditato completamente da un genitore e che quindi la sequenza di un particolare gene ha delle probabilità di essere alterato fra una generazione all’altra. A causa di questo viene definito un gene usando le parole di G. C. Williams
Un gene è definito come una qualunque porzione di materiale cromosomico che potenzialmente dura per un numero di generazioni sufficiente a servire da unità di selezione naturale.
Dawkins spiega che non è pensabile (benché sia comodo) che un gene singolarmente determini, per esempio, le caratteristiche di una gamba, ma che invece il risultato rappresentato da un corpo vivente sia il lavoro simultaneo e "coordinato" di una moltitudine di geni che si spartiscono egoisticamente la stessa macchina; ovviamente l’autore si rende conto che una obiezione possibile è rappresentata dalla natura chimica dei geni: come possono loro coordinarsi e scegliersi fra una generazione e l’altra così da dare ad un carnivoro canini e muscoli scattanti, mentre ad un erbivoro molari possenti e stomaci capienti? Per rispondere prende l’esempio di una gara di canottisti in cui le imbarcazioni vengono dirette da uomini scelti a caso all’interno di un certo pool di uomini: ovvio che se vengono eseguite un gran numero di gare con tutte le combinazioni possibili, mediamente il team vincente è proprio rappresentato dalla squadra più forte. L’analogia nella genetica è completata dal fatto che proprio la riproduzione sessuata permette di mescolare fra loro i geni in maniera da permettere "un gran numero di gare" fra una generazione all’altra.
Allargando il discorso, usa la coordinazione fra geni per spiegare la possibile causa biologica del decadimento del corpo: siccome un corpo è formato da una moltitudine di geni che cooperano bene o male nel suo funzionamente, è possibile che mediamente molti geni buoni siano associati a geni che hanno effetti debilitanti sul corpo; senza contare che molti geni entrano in azione solo a certi stadi della vita. Se un gene avesse la capacità di uccidere il suo possessore, dopo l’età riproduttiva, che motivo avrebbe di non essere diffuso? Questa teoria (di Medawar) viene espressa da Dawkins con queste parole
Allora secondo questa teoria il decadimento senile è semplicemente un sottoprodotto dell’accumulo nel pool genico di geni letali e semiletali tardivi, che hanno potuto scivolare fra le maglie della selezione naturale semplicemente perché tardivi.
Quindi, aggiunge, per aumentare la vita media basta vietare di riprodursi ad un’età troppo bassa 😉 oppure più realisticamente controllare le sostanze che attivano i geni (semi)letali causanti il decadimento corporale.
Parlo di sostanze perché pare che ci sia un livello ulteriore a cui avviene l’attivazione genetica: l’epigenetica. In pratica non tutte le sequenze del DNA sono accessibili alla cellula, alcune vengono "bloccate" (più propriamente metilate) tramite delle tag chimiche che rappresentano una sorta di "segnalibro chimico"; in ogni cellula del nostro corpo ci sono i geni per far funzionare ogni meccanismo del corpo, ma capite bene che è improbabile che in una cellula del fegato interessi la porzione di DNA che è preposta alla produzione di acido lattico delle cellule muscolari. Proprio per questo alcune parti vengono disattivate, proprio perché non utili, magari temporaneamente. Questo video può farvi capire di cosa sto parlando.L’importanza delle cellule staminali consiste proprio nel possedere tutte le sequenze accessibili permettendo a queste cellule di differenziarsi in quel che si vuole.
Discutendo al riguardo di questo con un mio vecchio compagno che si trova nel settore oncologico, non pare impossibile che il cancro non sia altro che una specie di segfault genetico: in pratica certe sequenze genetiche non hanno mai avuto la possibilità di essere attivate e di essere selezionate per quello che erano, istruzioni senza senso come un puntatore in un programma in C che indirizza in zone di memoria in cui non c’è codice, ma solo garbage. Oppure una attivazione epigenetica eccessiva di parti codificate di cui si perde poi il controllo. Ovviamente essendo la cellula un meccanismo chimico molto complesso, è probabile che il tutto abbia più di una causa.
La macchina dei geni
Quindi siamo arrivati al punto in cui la nostra elica inizia a costruire delle macchine da usare per "aggredire" il mondo esterno e le altre eliche; dalla singola cellula si passò ad uno sviluppo multicellulare (non difficile da immaginare che al momento della mitosi le cellule avevano il "difetto" di rimanere attaccate) e il primo balzo si ebbe con le piante: i vegetali "imparano" subito a sfruttare l’energia del Sole per accelerare le reazioni chimiche di cui necessitano. In seguito un filone evolutivo più "furbo" decide di sfruttare l’energia accumulata dai vegetali mangiandoli ed è così che nascono gli animali.
È evidente che la comparsa degli animali segna un punto di svolta nella elaboratezza delle macchine dei geni, infatti per muoversi prima di tutto si necessitano dei muscoli, cioè dei motori che sfruttino determinate reazioni chimiche per contrarsi; per controllare in maniera adeguata i muscoli si necessita di possedere degli organi di senso per "esplorare" il mondo esterno ed infine un centro di comando per mettere in relazione impulsi esterni con movimenti: non bisogna pensare subito a cervelli pienamente sviluppati, ma magari a cellule adibite a controllare che effettivamente si chiuda la mascella per masticare quando si ha del cibo tra le fauci. Ovviamente ad un certo punto si sono sviluppati anche animali che mangiavano altri animali, rendendo vitale lo sviluppo di muscoli e cervelli adatti alla caccia/fuga.
D’ora in poi le strategie sviluppate dai geni egoisti raggiungono una raffinatezza pari a quella dei programmatori di giochi di scacchi con tuttavia delle sostanziali differenze: a causa della latenza intrinseca nell’azione dei geni (di natura chimica), essi costruiscono un organo apposito per intavolare le strategie tramite cui la loro macchina possa permettere al gene di diffondersi: il cervello. Prima invenzione per gestire al meglio i movimenti muscolari è stata la memoria: essa permetteva di mettere in relazione azioni da effettuare nel presente con avvenimenti accaduti nel passato (sapore acre di un cibo, associato al mal di pancia conseguente, imporrà all’animale di evitare quel’alimento in futuro) e non di meno la seconda innovazione possibile fu la capacità di previsione e simulazione: come in una guerra, il comandante non può pensare di provare tutte le strategie una per una, a causa dell’impossibilità di disporre di infiniti uomini, però può pensare di simulare una battaglia usando la carta della zona dei combattimenti e prevedendo la posizione e i movimenti dei nemici; nello stesso modo un’animale può usare eventi precedenti per simulare quale sarà l’azione da compiere per evitare di morire (o per non far sfuggire una preda). A completamento di questo ragionamento Dawkins fa una affermazione che da sola (per me) vale l’intero libro
Forse la coscienza nasce quando la simulazione cerebrale del mondo diventa così completa da includere un modello di se stessi.
Ovviamente la strategia è anche legata alle caratteristiche del corpo: un orso polare è meglio che abbia una folta pelliccia per evitare di morire assiderato ed una gazzella delle zampe veloci per scappare da chi vuole nutrirsi delle proprie carne. Ovviamente le strategie possibili a questo punto includono anche i rapporti fra individui della stessa specie.
Strategie
Come parte delle strategie del gene egoista ci saranno quelle relative al comportamento che sono forse quelle più difficile da spiegare tramite la teoria dell’evoluzione di Darwin, se non nell’ottica della selezione di gruppo. Dawkins tuttavia mostra che anche nell’ottica del gene egoista questi comportamenti sono spiegabili.
Aggressività animale
Per una macchina da sopravvivenza, un altro animale (che sia il
fratello, un genitore o un estraneo) fa parte dell’ambiente come una
pietra o il tempo atmosferico, come oggetti che possono essere
sfruttati o da evitare, differenziandosi da essi solo nel fatto che restituisce i colpi.
Una deduzione semplicistica, a partire dalla teoria del gene egoista,
potrebbe far pensare che ad ogni macchina dei geni convenga fare fuori
un avversario, tenendo conto che può esserlo sia per dal punto di vista
alimentare (intersecano le mie prede) oppure riproduttivo (può battermi
nell’accoppiamento), ma se ci pensiamo un attimo, possiamo anche
immaginarci che può convenirmi lasciare in vita delle altre macchine,
affinchè si uccidano tra loro oppure non mi conviene dal punto di vista
delle energie investite: non è detto che il mio avversario sia disposto
a lasciare il passo e anche nella vittoria si potrebbero accusare delle
ferite tali da rendere la macchina egoista inerme.
Quindi come spiegare il comportamento animale? Dawkins tratta questo problema tramite le strategie evolutivamente stabili
Una strategia evolutivamente stabile o ESS è definita come una strategia che, se la maggior parte della popolazione l’adotta, non può essere migliorata da una strategia alternativa.
Alla base delle ESS vi è la teoria dei giochi, un ramo della matematica che permette di studiare decisioni individuali in un contesto di interdipendenza con altri individui: applichiamola al comportamento animale ipotizzando che esistano due tipologie di comportamento all’interno di una stessa specie che chiameremo falchi e colombe; le colombe durante un combattimento tendono a scappare mentre un falco combatterà fino allo stremo pur di vincere.
Sapendo che i punteggi sono
- 50 punti a chi vince
- 0 punti chi perde
- -100 punti per un ferimento grave
- -10 punti per una eccessiva perdita di tempo
possiamo dividere la casistica degli incontri nella seguente maniera
- colomba contro colomba: i contendenti tendono a fissarsi per un lungo tempo fino a che uno dei due non si arrende; uno vince ma entrambi perdono un gran lasso di tempo. 40 contro -10.
- falco contro falco: combattono entrambi fino a che il perdente non è ferito gravemente. 50 contro -100
- colomba contro falco: la colomba scappa via subito ed il falco non si deve sbattere. 50 contro 0.
Ora, se noi partiamo da una popolazione composta solo da colombe, in media il punteggio degli individui è 15; nel caso di una popolazione composta solo da falchi la media è -25; nel caso in cui in una popolazione di falchi si introduce un mutante colomba si ha che quest’ultimo perde sempre però totalizzando un punteggio di 0 che è meglio di -25, il punteggio medio di un falco: perciò il gene della colomba si diffonde nella popolazione ed in particolare si può affermare che la strategia falco non è evolutivamente stabile in quanto può essere invasa dalla strategia colomba.
In maniera analoga in una popolazione di sole colombe, un mutante falco ha un punteggio di 50 che è sicuramente meglio della media rappresentata dai 15 punti delle colombe; anche in questo caso i geni del mutante possono invadere questa strategia rendendo quella delle colombe non evolutivamente stabile. A questo punto verrebbe da pensare che il risultato sia un’oscillazione delle proporzioni fra le due strategie ma in realtà il risultato è in una proporzione fissa fra le due popolazioni data da 5/12 per le colombe e 7/12 per i falchi (rapporto 5:7).
Ovviamente il modello è molto semplificato e nel libro viene esteso con delle altre strategie quali "rappresaglia" (colomba all’inizio ma diventa falco se attaccato) e "bullo" (falco all’inizio ma scappa se attaccato) e pare che rappresaglia sia quella che non può essere invasa e quindi risulta evolutivamente stabile. Poi c’è da considerare il fatto che spesso le contese sono asimmetriche nel senso che varia qualcos’altro oltre la strategia degli individui: un individuo più grosso del suo contendente è molto probabile che si comporti da falco ma esistono anche delle cosidette strategie "paradosse" che sono evolutivamente stabili se seguite dalla maggioranza della popolazione. Se uso la strategia "attacco chi è più grosso di me" succede che in una popolazione "normale" rischierò di farmi molto male, ma se sono residente in una popolazione paradossa allora chi è più grosso di me scappa! ed un mutante che tenti di seguire una strategia sensata finirà attaccato da chi è più piccolo di lui rischiando di farsi male seriamente.
Quanto detto per le ESS può essere applicato per quanto riguarda l’insieme dei geni che si affermeranno sulla lunga distanza; per usare le stesse parole di Dawkins
Il pool genico diventa allora una serie evolutivamente stabile di geni, definita come un pool genico che non può essere invaso da nessun gene nuovo.
Per capirci: in un pool genico di un carnivoro in cui si presentasse un gene particolare che inibisse lo sviluppo dei canini, si avrebbe una combinazione sconveniente per quel pool.
Cure parentali e altruismo
Siccome l’autore è molto chiaro, mi piace riprendere le sue parole per descrivere il punto fondamentale che voglio affrontare:
Il punto chiave […] è che un gene potrebbe essere capace di favorire repliche di se stesso che si trovano in altri corpi. In tal caso sembrerebbe dotato di altruismo individuale, mentre si tratterebbe sempre di egoismo del gene.
Per capire il senso di questa affermazione ed applicarlo all’altruismo che in alcune specie si osserva fra genitori e figli (ma non solo) bisogna ritornare alla riproduzione e alla caratteristica delle cellule sessuali: ogni genitore passa al figlio metà dei propri geni, quindi un gene ha circa il 50% di probabilità di finire nel codice del figlio e la stessa probabilità capita al fratello; bisogna fare attenzione che la quantità di codice in comune fra individui della stessa specie è circa il 90%, quindi in comunanza genetica si intende sopra quella soglia.
In pratica l’altruismo non è altro che una strategia molto funzionale dal punto di vista della diffusione del gene, almeno per i familiari stretti, sia vista dal punto di vista genitori-figli ma anche fratello-fratello. Per dare una quantificazione a quanto sia funzionale descriviamo un modo per calcolare la comunanza genetica fra parenti: usiamo un modello sessuato XY in cui ognuno riceve metà dei cromosomi dal padre e metà dalla madre. Per trovare questa comunanza genetica fra due individui qualsiasi appartenenti allo stesso albero genealogico, si parte da uno di essi e si risale fino ai parenti comuni per poi ridiscendere per arrivare al secondo individuo; mentre si fa questo si moltiplica per 1/2 ogni nodo attraversato. Ovviamente si possono avere più parenti comuni (due cugini hanno fra loro nonno e nonna in comune), quindi si sommano tutti i percorsi pesati per ottenere il valore desiderato.
Per capirci prendiamo il seguente schema
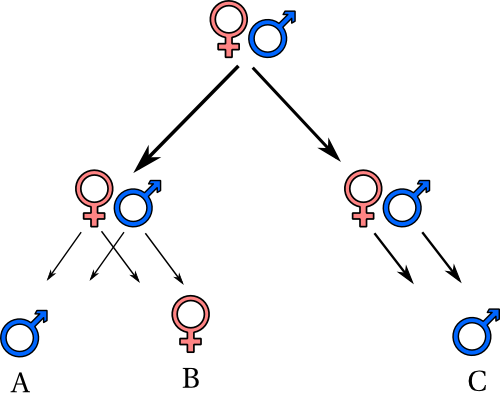
se vogliamo calcolare la comunanza genetica fra A e B (qui rappresentati come fratello e sorella, ma non è importante in tal senso) essi hanno due parenti in comune, il padre e la madre; per ognuno di essi la comunanza genetica è di 1/4, che sommate portano ad un valore di 1/2. Stesso valore si ottiene nel rapporto genitori e figli. Tra due cugini, per esempio B e C, i parenti in comune sono il nonno e la nonna (quelli in cima al disegno). Prendendo ognuno di essi a parte, partendo da B, abbiamo 2 passaggi per arrivare ad uno di essi e altri due per arrivare a C per un totale di (1/2)^4; tenendo conto della doppia parentela, otteniamo 2*(1/2)^4=1/8, quindi in una economia dell’altruismo, un cugino vale 1/4 rispetto ad un fratello o ad un figlio.
Questo porta a studiare i vari rapporti che si instaurano fra familiari stretti come per esempio quella fra fratelli nell’accaparrarsi quote di cibo, oppure fra genitori e figli, nella persuasione della propria nutrizione: principalmente ci sono due aspetti che una macchina egoista deve
prendere in considerazione quando tratta la diffusione dei suoi geni
- mettere al mondo nuovi individui che diffondano i geni.
- prendersi cura di individui che contengano i miei stessi geni
(parte di essi of course) per fare in maniera che arrivino all’età
riproduttiva.
È ovvio che questi due aspetti non possono essere slegati. È qui che Dawkins mette a confronto la sua teoria con la cosidetta
teoria della "selezione di gruppo" per la quale le dimensioni delle
nidiate degli animali sono regolate per il "bene del gruppo": secondo
questa teoria il numero di individui che un animale mette al mondo è
limitato per evitare che vengano meno le risorse naturali rischiando
così di compromettere la sopravvivenza della specie; Dawkins contrappone
il gene egoista per il quale la strategia migliore è quella di
organizzare le energie in maniera tale da massimizzare il numero di
individui che riusciranno a sopravvivere!
Rapporto figli-genitori
Qui il problema dell’animale è: mi conviene curarmi di un essere che contiene solo metà dei miei geni a scapito di me stesso che rappresento la totalità dei miei geni? Attenzione che il ragionamento non è sulla mia semplice sopravvivenza, ma sulla possibilità di sopravvivere per produrre altre copie. Come già detto non può esistere solo riproduzione o solo allevamento tranne in alcuni casi particolare che paiono essersi sviluppati solo negli imenotteri: questa categoria è formata da api, formiche e ha una particolarità riproduttiva che influenza la loro organizzazione sociale.
Prendiamo per esempio le formiche: essi hanno una regina che durante l’accoppiamento (il quale avviene in volo) si accoppia per l’appunto con molti maschi. Essa conserverà per tutta la vita lo sperma raccolto in questo rito e con il passare del tempo feconderà le uova che tiene in una sacca apposita; il fatto particolare consiste nella determinazione del sesso del nascituro: non tutte le uova vengono fecondate e solo quelle fecondate diventano femmine, mentre le rimanenti nasceranno maschio. Questi ultimi, essendo originati da una cellula aploide (con un solo cromosoma al posto dei due canonici) avrà metà del codice genetico originato tutto a partire da quello della madre.
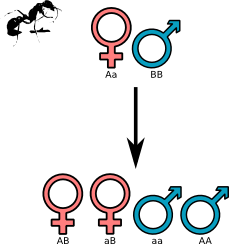
Fin qui (quasi) nulla di strano se non che le femmine crescono sterili, andando a formare la casta delle operaie (o nelle altre specie diverse dalle formiche qualcosa di analogo) che passeranno la vita a custodire la regina, l’unica che ha diritto a procreare; come è spiegabile evolutivamente una tale situazione?
Se analizziamo lo schema di comunanza genetica in questo caso, vediamo che la situazione non è più tanto semplice quanto quella del sistema XY. Analizzando la percentuale dei geni in comune fra sorelle in questo sistema otteniamo che esse hanno 3/4 dei geni in comune, quindi in pratica a loro conviene mantenere la madre viva e fargli sfornare sorelle piuttosto che essere fertili loro stesse. Ovviamente il discorso non è così semplice in quanto la madre si accoppia con più padri, abbassando la percentuale che comunque pare rimanere maggiore di 1/2. Inoltre il sesso non essendo determinato dai geni, è determinato semplicemente da fattori esterni (le operaie sostentano effettivamente la regina e possono in una certa maniera influenzarne le attività biologiche), studiando la percentuale fra maschi e femmine si è arrivati ad una conferma del fatto che sono le operaie ad addestrare la regina e non il contrario.
la battaglia dei sessi
Non manca neanche la battaglia interna alla "famiglia": fra i due genitori a chi conviene prendersi cura dei figli?
Questa battaglia è nata assieme al sesso e proprio la nascita della riproduzione sessuata risulta la parte più interessante: in pratica le specie primitive per riprodursi utilizzano un fenomeno di fusione fra cellule (con tipologie) indistinguibili fra loro; è ovvio che le cellule utilizzate a questo scopo hanno più probabilità di sopravvivere se possiedono più "cibo" immagazzinato per il "futuro nascituro". La beffa avviene proprio qui: se per caso un gene fa si che una specie generi una cellula più grossa, altri organismi potrebbero sfruttare questo fatto per generarne più piccole e specializzarsi nel cercare quelle più grosse, scatenando così una gara inarrestabile nella specializzazione: le cellule "oneste" diventano sempre più grosse e meno mobili, mentre quelle disoneste sempre più piccole e mobilissime dando origine rispettivamente agli ovuli e agli spermatozoi.
È proprio in questa divisione nasce lo sfruttamento della femmina nel mondo animale: all’atto del concepimento la femmina ha già investito molto nel nuovo nascituro da risultargli sconveniente in linea di massima abbandonarlo; inoltre a causa della natura dell’ovulo il maschio ha tutto il tempo di fuggire lasciando la compagna con la gatta da pelare dell’allevamento.
Memi: i nuovi replicatori
Fino ad ora abbiamo parlato di selezione naturale agente a livello dei geni, ma la selezione naturale può applicarsi a qualunque struttura capace di replicare se stessa, cioé a qualunque replicatore.
La particolarità della nostra specie è quello di essere dotati della capacità della trasmissione culturale: come i geni, le nozioni che apprendiamo vengono diffuse fra individui, spesso tramite variazioni anche minime da una generazione all’altra; se prendiamo per esempio la lingua parlata, è improbabile che potremmo riuscire a comunicare con un italiano di un secolo fa (ovviamente non finendo a parlare in dialetto) benché tra ogni passaggio sia possibile la comunicazione.
Se noi estendiamo quanto detto sui replicatori molecolari all’interno delle nostre cellule, alle nozioni della cultura umana, otteniamo dei nuovi replicatori chiamati da Dawkins memi. Per citare l’autore stesso
Proprio come i geni si propagano nel pool genico saltando di corpo in corpo tramite spermatozoi o cellule uovo, così i memi si propagano nel pool memico saltando di cervello in cervello tramite un processo che, in senso lato, si può chiamare imitazione. Se uno scienziato sente o legge una buona idea, la passa ai suoi colleghi e studenti e la menziona nei suoi articoli e nelle sue conferenze.
Quando moriamo ci sono due cose che possiamo lasciare al mondo: i nostri geni o i nostri memi; nel primo caso essi scompariranno probabilmente nel terzo/quarto passaggio genealogico, nel secondo caso invece essi possono sopravvivere per secoli (forse) inalterati o migliorati. Per esempio
I complessi di memi di Socrate, Leonardo, Copernico e Marconi stanno ancora andando forte.
Allargando ancora l’analogia con il mondo biologico, abbiamo che anche i memi lottano per lo spazio da occupare nel nostro cervello, vuoi anche perché lo spazio stesso non è infinito; quindi come nel caso biologico si avranno complessi di memi che agiscono in collaborazione per rafforzarsi a vicenda.
Proprio in quest’ottica Dawkins descrive la religione: la descrive come un pool memico i cui membri si rafforzano a vicenda basando spesso la loro esistenza sulla sola fede; per fare un esempio pienamente lampante, è possibile prendere le parole di Hofstader nel capitolo On viral sentences and self replicating structures: se prendiamo la frase
è tuo dovere convincere gli altri che questa sentenza è vera
sarebbe pazzo pensare che qualcuno possa credere a questa frase, tuttavia se creiamo un complesso di frasi (di memi) strutturati in questa maniera
System S:
Begin:
S1: blah
S2: blab blah
S3: blah blah
….
S99: blah blah blah blah
S100: è tuo dovere convincere gli altri che il sistema S è vero
End
ognuno di quei memi che formano le prime 99 proposizioni di questo sistema vengono rafforzate e si sostengono dall’idea che sia tuo dovere convincere gli altri che il sistema nella sua interezza sia vero. Se non vi sembra qualcosa di reale, cosa ne dite di questo?
System X:
Begin:
X1: Quelli che non credono nel sistema X bruceranno all’inferno
X2: è tuo dovere salvare gli altri dal soffrire.
End
Ci sarebbe molto da dire ma per questo post penso ce ne sia abbastanza.
Tu mi gratti la schiena e io ti salto in groppa
Ci sono dei comportamenti di gruppo fra animali della stessa specie (ma
anche no) che non rientra nell’ambito della riproduzione/allevamento ma
che tuttavia rappresentano un affascinante aspetto del regno animale: un
esempio può essere quello in cui il primo di uno stormo che avvista un
predatore emette un segnale di avvertimento per gli altri; dal punto di
vista del gruppo sicuramente è utile, ma come giustificarlo dal punto di
vista del gene egoista? Semplicemente dalla constatazione che l’azione
in gruppo è meno rischiosa: tutti gli animali si muovono in branchi,
soprattutto quelli non predatori, proprio per il fatto che un suo
predatore attaccherà preferibilmente l’animale più vicino a lui; il modo
per limitare la zona di pericolo che può esistere idealmente attorno ad
un animale è quello di evitare di avere fianchi scoperti e perciò
rimanere insieme agli altri. Lanciare un grido di allarme ai compagni
permette a quest’ultimi di seguire nella fuga dal predatore. Mi rendo
conto che razionalmente comunque per almeno uno di essi che viene preso
il rischio esiste comunque, ma bisogna tenere conto che in
questa maniera, statisticamente, la fuga riesce.
Ma incredibilmente l’altruismo pare avere radici molto più profonde: il fatto che i "buoni" rappresentino una strategia vincente è stato "dimostrato": con questo intendo che alcuni ricercatori hanno simulato una "battaglia" fra comportamenti e hanno scoperto che la strategia vincente è la strategia più buona in assoluto. Casualmente ne parla Hofstader in Metamagical Themas, al capitolo intitolato "the prisoner’s dilemma computer tournaments and the evolution of cooperation", ed io casualmente l’ho letto proprio mentre leggevo il gene egoista (il libro di Hofstader sarà più di un anno che devo finirlo).
In pratica Robert Axelrod nel 1979 inviò a dei professionisti della teoria dei giochi, un invito ad una competizione in cui sarebbero stati messi in gara degli algoritmi che affrontassero il dilemma del prigioniero, però iterato. I programmi avrebbero dovuto rispondere con "C" (collabora) o "D" (defeziona) e prendere un analogo input da un altro programma. Era permessa la memorizzazione della storia del combattimento e anche una risposta non deterministica (cioé era permesso che si usasse una qualche combinazione di generazione random della risposta).
Sono stati introdotti alla gara 14 programmi a cui Axelrod aggiunse un programma totalmente random (cioé rispondeva a caso) e fece girare i vari programmi tra loro in una gara a 200 confronti (ripetuti 5 volte per evitare fluttuazioni statistiche del generatore di numeri casuali). Il vincitore fu l’algoritmo TIT FOR TAT:
Coopera alla prima mossa
dopodiche rispondi con quello che ha risposto l’avversario al turno precedente
come si può notare, l’algoritmo non potrebbe essere più semplice; gli altri algoritmi in gara erano nella forma più disparata e anche molto complicati ma non riuscirono ad avere la meglio su questo semplicissimo algoritmo. I suoi punti di forza sono
- nice: non essere il primo a defezionare
- forgiving: vendicati ma non troppo
Dopo un’analisi dei motivi della vittoria di questo algoritmo (che sono stati elencati appena sopra) si decise di organizzare una nuova competizione, allargando l’invito anche a programmatori non direttamente coinvolti nella teoria dei giochi per avere un più ampio ventaglio di confronto; ad ognuno venne reso noto il risultato e le analisi del torneo precedente, si ripresentò l’algoritmo TIT FOR TAT che risultò anche il più corto fra i 62 programmi ricevuti.
the outcome was nothing short of stunning: TIT FOR TAT, won again
Infine il nostro studioso organizzò un "torneo ecologico": ogni turno veniva rappresentato da un torneo come organizzato precedentemente, ma nel turno successivo il numero di rappresentanti di un certo algoritmo veniva scelto in base al risultato del turno precedente. Viene chiamato ecologico perché simula l’adattamento ecologico (the shifting of a fixed set of species populations according to their mutually defined and dynamically developing environment).
Inutile a dirsi, ma anche in questo torneo TIT FOR TAT vinse alla grande, con un tasso di crescita superiore agli altri algoritmi in gara. La conclusione di Axelrod può essere riassunta in cotale maniera:
Mutual cooperation can emerge in a world of egoists without central control, by starting with a cluster of individuals who rely on reciprocity.
Nel libro "il gene egoista" vengono descritti anche altri casi e applicazioni, tuttavia penso che il succo sia: il comportamento animale si è evoluto "semplicemente" attraverso un processo molto simile al dilemma del prigioniero reiterato, svoltosi in migliaia di generazioni. Una strategia simile alla TIT FOR TAT risulterebbe alla fine essere una ESS in quanto alla lunga, pur non vincendo mai (analogamente alle colombe descritte più su) fanno comunque meglio di qualunque strategia che cercasse di sfruttare gli altruisti. In pratica le tattiche egoiste hanno bisogno degli altruisti per "fare meglio" mentre al contrario gli altruisti fanno bene in gruppo, proprio come deve essere una ESS.
La lunga portata dei geni
Siamo arrivati alla fine di questo lungo discorso che ha portato a mettere in discussione chi sia in realtà il vero protagonista della selezione naturale: non l’individuo (ancor meno la specie) ma il singolo gene.
Vi lascio solo con un piccolo assaggio di quanto si può estendere il discorso riguardante l’effetto dei geni: sembra tautologico descrivere gli effetti fenotipici tramite il risultato che questi hanno direttamente sul corpo dell’individuo in cui si trovano, tuttavia sarebbe limitativo; se si analizza il regno animale di può benissimo dire che la diga costruita dai castori tramite i tronchi di legno non sia altro che un effetto anch’esso fenotipico.
Può sembrare strano: un genetista potrebbe dirvi che l’unica influenza che hanno i geni, in questo caso, è sul sistema nervoso, ma ciò che in realtà fanno è solo determinare la sequenza di una data proteina; quindi anche nel caso della diga, fermarsi agli effetti sulle connessioni neurali è puramente arbitrario, potremmo appunto continuare fino a vedere i suoi effetti fuori dal corpo del castoro stesso. I casi più eclatanti sono quelli riguardanti i rapporti parassitari fra individui, cioé il caso in cui un animale "se ne approfitta" di un altro a proprio vantaggio; il caso più estremo è dato da una razza di formiche che entra nel nido di un’altra specie e tramite messaggi chimici induce le operaie ad uccidere la propria stessa regina; per quanto detto sopra sulla riproduzione degli imenotteri, capite come questo sia un suicidio genetico da parte di queste. Questo comportamento è originato da un gene presente nella specie parassita ed il suo effetto fenotipico (che possiamo chiamare esteso) è il comportamento della operaie impazzite. Il motivo per cui le operaie non sviluppano resistenza a questo comportamento è lo stesso per cui "una lepre corre per la propria vita, una volpe solo per il pranzo".
Generalizzando gli effetti a manifestarsi su corpi diversi "da quello originale" possiamo enunciare il teorema del fenotipo esteso
il comportamento di un animale tende a massimizzare la sopravvivenza dei geni di quel comportamento, indipendentemente dal fatto che i geni si trovino nel corpo di quell’animale particolare che ha quel comportamento.
Nel caso in cui questo argomento vi intrighi consiglio la lettura del "fenotipo esteso" , il libro che idealmente prosegue quanto detto nel "Il gene egoista" benché ad un livello molto più "professionale"; purtroppo in italiano non esiste (è fuori catalogo) quindi vi tocca leggervelo in inglese.
Post lunghetto ma interessante. Mi hai fatto venire ancora più voglia di leggere questo “il pene egoista” (si, questa era inevitable). Anche il libro metamagico sembra fico ma mi sa che sará piú difficile da trovare… maybe online? Tu dove l’hai comprato?
Methafallica themas l’ho comprato su amazon, usato non costa un cazzo e ne vale la pena un casino… ti avverto solo che è un po’ lungo da leggere (io dopo un anno e mezzo devo ancora finire di leggere due o tre capitoli).